检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
/ 澳门太阳集团官网www / 学术科研
2021年5月21日,我院邓宏魁研究组在Nature Communications杂志上发表了题为“Chemically Defined and Xeno-free Culture Condition for Human Extended Pluripotent Stem Cells”的研究论文,在体外建立了成分明确、无异源成分的人潜能扩展多能干细胞(EPS细胞)的培养体系,并利用该体系建立了将体细胞诱导为无异源人EPS细胞的重编程诱导方法。
如何建立高质量的多能干细胞一直是干细胞研究的核心问题。EPS细胞是2017年邓宏魁研究组建立的一类具有全能性功能特征的多能干细胞,同时具有胚内和胚外组织的发育潜能(Yang et al., 2017)。自建立以来,EPS细胞得到了广泛的应用。邓宏魁研究组利用EPS细胞建立了快速制备基因修饰动物模型的技术(Li et al., 2019a),并利用人EPS细胞高效制备功能性肝细胞(EPS-Heps)(Wang et al., 2020),与传统人多能干细胞制备的肝细胞相比,EPS-Heps在转录谱上更类似于人原代肝细胞。该研究结果突出了EPS细胞产生功能性细胞的应用潜力,可以为体外制备功能细胞提供更好的来源。国际其他研究组也在EPS细胞的研究和应用中取得了一系列重要进展,利用 EPS细胞在体外构建功能性的早期胚胎结构(Li et al., 2019b; Sozen et al., 2019);将人EPS细胞注射到食蟹猴早期囊胚中,制备了人猴嵌合体胚胎,并成功使胚胎存活时间达到20天,这是目前猴子胚胎体外培养能达到的最长时长(Tan et al., 2021)。这些研究都表明EPS细胞具有广泛的应用前景和优势。但是,人EPS细胞的培养依赖于饲养层细胞,制约了EPS细胞的进一步转化应用。
为了解决这一问题,邓宏魁研究组通过对饲养层分泌的细胞因子和胞外基质等进行研究,建立了成分确定的,无异源的培养体系(xeno-free EPS culture condition,图1)。无饲养层、无异源体系培养的人EPS细胞能够在体外长期稳定地培养60代以上,且能维持人EPS细胞的基本性质。转录组分析表明,无异源人EPS细胞具有和饲养层细胞培养的人EPS细胞相似的整体表达谱,与EPS特性相关的早期胚胎富集的基因模块在无异源培养的人EPS细胞同样富集。而且,利用该培养体系与重编程技术结合能高效建立人体细胞来源的无异源成分的人EPS细胞系,为制备人EPS细胞提供了新的方法。
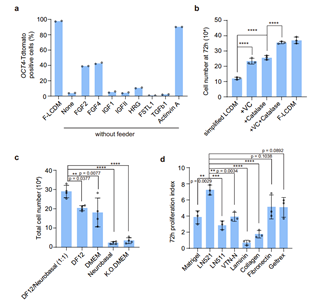
图1:成分确定的、无异源的人EPS细胞培养体系的建立
邓宏魁研究组进一步系统研究了无异源培养的人EPS细胞的胚外发育潜能,将其注射到早期小鼠胚胎后,能够稳定地嵌合到小鼠胚胎期E6.5天的胚内和胚外组织。进一步的实验发现,无异源的人EPS细胞若培养在滋养层干细胞培养基中,能够转化成滋养层干细胞类似性质的细胞;ATAC-seq测序结果发现在无异源的人EPS细胞中,与胚外发育相关的基因位点处于更开放的状态(图2)。这些结果都证明无异源培养的人EPS细胞具备胚外组织的发育潜能。
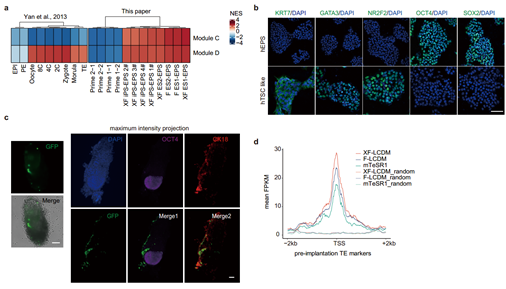
图2:xeno-free hEPS细胞的胚外发育潜能
该培养体系排除了饲养层细胞不确定成分的影响,提高了人EPS细胞培养的稳定性,为推动人EPS细胞在分化,疾病模型和发育生物学的广泛应用提供了条件。无异源成分的人EPS细胞培养体系为建立符合临床应用标准的人EPS细胞系奠定了技术基础,未来将进一步推进EPS技术向临床的转化应用。
太阳集团见好就收9728邓宏魁教授、太阳集团见好就收9728干细胞研究中心徐君研究员和白云教授为该论文的共同通讯作者,太阳集团见好就收9728李程研究员为该论文的生物信息学分析提供了重要支持,刘蓓博士,博士研究生陈诗,徐亚星博士,博士研究生吕钰麟为该论文的共同第一作者。本工作获得了国家重点研发计划、国家自然科学基金、北京市科委的资助。
原文链接:https://www.nature.com/articles/s41467-021-23320-8
参考文献
Li, H., Zhao, C., Xu, J., Xu, Y., Cheng, C., Liu, Y., Wang, T., Du, Y., Xie, L., and Zhao, J. (2019a). Rapid generation of gene-targeted EPS-derived mouse models through tetraploid complementation. Protein & cell 10, 20-30.
Li, R., Zhong, C., Yu, Y., Liu, H., Sakurai, M., Yu, L., Min, Z., Shi, L., Wei, Y., and Takahashi, Y. (2019b). Generation of blastocyst-like structures from mouse embryonic and adult cell cultures. Cell 179, 687-702. e618.
Sozen, B., Cox, A.L., De Jonghe, J., Bao, M., Hollfelder, F., Glover, D.M., and Zernicka-Goetz, M. (2019). Self-organization of mouse stem cells into an extended potential blastoid. Developmental cell 51, 698-712. e698.
Tan, T., Wu, J., Si, C., Dai, S., Zhang, Y., Sun, N., Zhang, E., Shao, H., Si, W., and Yang, P. (2021). Chimeric contribution of human extended pluripotent stem cells to monkey embryos ex vivo. Cell 184, 2020-2032. e2014.
Wang, Q., Sun, D., Liang, Z., Wang, J., Zhong, X., Lyu, Y., Cao, J., Lin, Z., Du, Y., and Miao, Z. (2020). Generation of human hepatocytes from extended pluripotent stem cells. Cell research 30, 810-813.
Yang, Y., Liu, B., Xu, J., Wang, J., Wu, J., Shi, C., Xu, Y., Dong, J., Wang, C., and Lai, W. (2017). Derivation of pluripotent stem cells with in vivo embryonic and extraembryonic potency. Cell 169, 243-257. e225.
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号